Locomotory and Feeding Ecology:
Copepods and Friends
Shelley Rosenthal1
Fall 1996
Abstract
Locomotion and feeding processes are of great importance when considering the ecology of organisms. Animals are adapted to their fluid environment. Copepods are studied as an example of locomotory and feeding mechanisms in water. The Reynolds number was calculated for a swimming copepod, and indicates that inertial and laminar flow were present. Feeding behavior is examined in different algal concentrations, and grazing rate is determined.
Introduction
The processes of feeding and locomotion in the world are key to the ecology of all organisms. There is a multitude of methods by which organisms move, and gather food. Bipedal walking in humans, flying insects, and swimming fish, all, indeed, occur in a fluid environment. Organisms are morphologically adapted to their surrounding medium, be it air or water. The effectiveness of such adaptations, is very much determined by the scale at which these processes are occurring. For example, comb-like appendages at low Reynolds numbers may be functionally comparable to paddle-like appendage at high Reynolds numbers (Vogel, 1994). Reynolds numbers represent the ratio of inertial to viscous forces (Ligget, 1994), and determine the behavior of flow around an organism. Large animals, due to their large size and high relative velocity, exist at high Reynolds numbers; while small animals, due to their small size and relatively low velocities, exist at low Reynolds numbers. A large animal must therefore exert large amounts of energy to initiate motion and overcome the inertial force, but is also carried forward by the same force after swimming stops (Brusca and Brusca, 1990). A small animal would expend less energy to start motion, but would stop immediately after exertion ceased.
There is great diversity among the locomotory appendages and processes of different organisms. Animals that swim generally do so through ciliary and flagellar motion (i.e. veliger larvae), jet propulsion (i.e. cnidarians), or through tail propulsion (i.e. fish). Swimming organisms, however, are not the only ones that must cope with life in flow. Animals that fly are also at the mercy of fluid dynamic forces. Flying animals may hover, soar, or employ powered flight.
Suspension feeders may be of two types: those that rely on current to provide a never ending supply of nutrients, and those that generate their own feeding currents. The former being passive suspension feeders, and the latter being active suspension feeders. While ‘suspension feeding’ refers mainly to the nature of location of food, ‘filter feeding’ refers to how the particulate nutrients are captured (Vogel, 1994). Filter feeding may occur through many capture mechanisms, including direct interception, inertial impaction, gravitational deposition, motile-particle deposition, and electrostatic attraction (Rubenstein and Koehl, 1977). Feeding on suspended particulate matter, therefore, is not limited to sieving mechanisms alone.
Most of the animals in the class Copepoda are marine, and motile, feeding on phytoplankton (Pechenik, 1996). They normally range in size from 0.5-5.0 mm, with a few large, parasitic species (Brusca and Brusca, 1990). Feeding is accomplished through currents generated by the antennae and mouth. The second maxillae capture particles without contact, drawing food filled fluids in to the interlimb space. It is the maxillae that then transport the food to the area of capture, with no filtering ever involved (Stricker, 1982). Diel rhythms are know to exist in the feeding behavior of copepods (Duval and Geen, 1976;Conover et al., 1986), as well as discriminate selection between algae of different nutritional content (Cowles et al., 1988).
Materials and Methods:
Locomotion
A sample of calaniod copepods was collected from Shane Lake, British Columbia. The sample was stored in an aerated container of lakewater, under normal laboratory conditions. The experiment occurred from 1800-2100h, again under normal laboratory conditions. Several copepods were placed in a single Petri dish, and observed under a dissecting microscope. A ruler, marked in 1mm increments, was used to determine the average length of the animal. The size of the field of view was measured, and the time take for a copepod to swim the distance was recorded for each of the five trials. The animals length and velocity were then used to calculate the Reynolds numbers for the swimming animal, using the formula Re = ul/v.
Feeding
A known volume of algal (Chlorella pyrenoidosa) stock solution and water from the Torpy River system was prepared for a total volume of 00mL. Concentrations were labeled as belonging to group i=1-4, with Ci = 100 cells/mm3, 80 cells/mm3, 370 cells/mm3, and 200 cells/mm3, respectively. Twelve copepods were placed in each of four solutions, of varying concentrations. A control solution was prepared, with the same concentration as the most dilute of the experimental concentrations. This solution was used as an index of settling, and subtracted from the grazing rate to separate animal activity from settling. The algal cell concentration was measured at time intervals (0 min, 15 min, 30 min, 45 min, and 60 min), using a Ward’s haemocytometer. Due to time constraints, each solution was monitored by a different group of students.
Results
1. Locomotion
The copepod measured was approximately 2 mm in length. While under observation,
the copepod appeared to exhibit two types of swimming behavior. The first was slow motion, and changes in direction were common, while the second appeared to be much faster. When measuring the time taken to cross the field of view, it was the average of the two types of motion that was measured. The velocity ranged from 0.018 ms-1 to 0.045 ms-1 (mean = 0.027 ms-1, s = 0.010), giving Reynolds numbers from 35-39 (mean = 53, s = 20). These results indicate inertial conditions and laminar flow.
Feeding
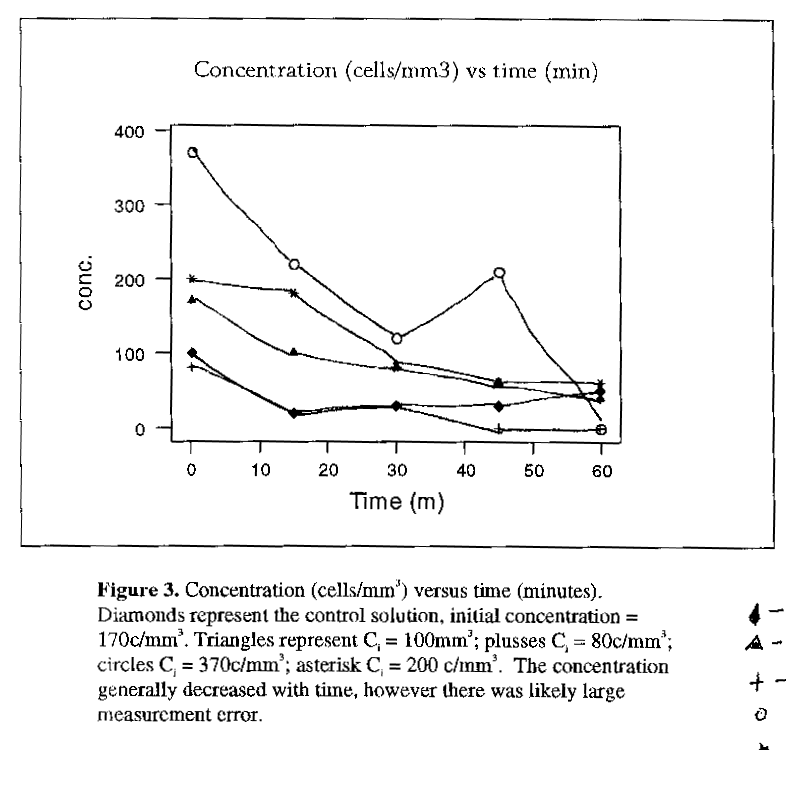
While the algal cell concentrations generally decreased with time (Fig. 3), it is not possible to draw any conclusions from the data (appendix 1). An ANOVA showed that there was no difference in algal concentrations between times in groups 1-3, due to high variances (group 1 F(4, 20) = 2.34, p =0.09, group 2 F(4, 20) = 1.46, p = 0.25; group 3 F(3, 16) = 2.37, p = 0.11). The control and group 4 demonstrated change in algal cell concentrations through time (group 4 F(4, 20) = 5.51, p = 0.004; control F(4,20) = 3.33, p = 0.03). Standard deviations of the cell counts were often greater than the means, indicating large measurement error. As, well, the concentrations increased over some of the time intervals, impossible, considering there was no input of algal cells, and two methods of loss (copepod feeding, and settling). The calculated grazing rate for the control solution was used to indicate settling, but for some of the time intervals, the rate was larger than the measured cell loss, including both settling and grazing (see Appendix 2).
Discussion
Organisms exhibit a wide variety of adaptations to aquatic habitats. Adaptations to avoid sinking include swimbladders, ammonium-rich body fluids, and eddy support. Copepods alternate between swimming in short bursts (0.3 ms-1), and sinking (Alexander, 1990). Fish are made efficient swimmers due to their control of vortices (Triantafyllou and Triantafyllou, 1995), and may be specifically adapted for cruising, accelerating, or maneuvering (Webb, 1992). The ‘stop and start’ type swimming observed cannot therefore be attributed to high viscosity, because the calculated Reynolds numbers indicate inertial conditions (Re>1).
The feeding trial did not reveal any significant trends in grazing rate. While the tendency was for concentration to decrease with time, this was only significant for the control (adj. r2 = 86.7%, p = 0.014), and for concentration 4 (adj. r2 = 80.6%, p = 0.025). A large amount of settling occurred (as seen by the grazing rates for the control solution, Appendix 2). The lack of significant findings was likely due to the large measurement error. As well, the experiment was carried out in full light. Had there been a diel pattern to the vertical distribution of the copepods in Shane Lake, they would not have been feeding under such conditions. The shock of a new thermal and solute environment may have been detrimental to the animal’s health. The animals may not then be feeding, even though they likely had not fed for several days.
Literature Cited
Alexander, McNeill R. 1990. Size, speed and buoyancy adaptations in aquatic animals. Amer. Zool. 30:189-196.
Brusca Richard C., and Gary J. Brusca. 1990. Invertebrates. Sinauer Associates Inc.
Sutherland, Massachusetts.
Conover, R.J., A.W. Herman, S.J. Prinsenberg, and L.R. Harris. 1986. Distribution
of and fedding by the copepod Pseudocalanus under fast ice during the arctic
spring. Science 251:1245-1247.
Cowles, T. J., R. J. Olson, and S.W. Chisholm. 1988. Copepod feeding. Marine
Biol. 100: 41.
Duval, W.S. and G.H. Geen. 1976. Diel feeding and respiration rhythms in
zooplankton. Limnol. Oceanogr. 21(6):823-828.
Ligget, James A. 1994. Fluid Mechanics. McGraw-Hill Inc. Toronto.
Pechenik, Jan A. 1996. Biology of the Invertebrates, 3rd ed. Wm C. Brown
Publishers, Toronto.
Rubenstein, Daniel I., and M.A. R. Koehl. 1997. The emchanisms of filter feeding:
some theoretical conditions. Am. Nat. 111(981):981-993.
Strickler, J. Rudi. 1982 Calanoid copepods, feeding currents and the role of gravity.
Science 218:158-160.
Triantafyllou, Michael S., and George S. Triantafyllou. 1995. Sci. Amer. March p64-
70.
Vogel, Steven. 1994. Life in Moving Fluids: the physical biology of flow. Princeton
University Press. Princeton, New Jersey.
Webb, Paul W. 1994. Form and function in fish swimming. Sci. Amer. July p72-
82.